【原创】真核细胞起源的科学猜想 — 兼融假说
河南科技大学 食品与生物工程学院 WLG团队
(导师:孙军杰;团队成员:王德保、刘美强、王小磊、郭叶青)
摘 要 :真核细胞的起源是生物进化过程中重要的环节,在生物进化中有着举足轻重的地位。对真核细胞起源的研究,不仅有利于解释远古时代各种各样奇异的生命进化现象,更有利于我们对人类的起源有一个更为深刻的认识。随着科学技术水平的提高,生物学也蓬勃发展起来,对真核细胞起源也有了更进一步的认识,各种各样与真核细胞起源有关的学说也不断地被发表出来。这也为我们的研究工作奠定了一个坚实的基础,我们WLG团队将前人的研究成果有机的结合了起来,主要的成果有:病毒起源的内源性学说,内共生与非内共生学说,生物界三域分类学说,古核生物与真核生物、原核生物的研究,寒武纪生命大爆炸与二叠纪生命大灭绝的研究等,在此基础上,我们大胆猜想,并做了更近一步的探究,为此,我们提出了更为科学的猜想——兼融假说(图1),并提出生物界四域分类学说(图2)。兼融假说融合了前辈们各种假说的优点,弥补了各种假说的不足,不仅解释了原始病毒的起源、内含子的起源、细胞核的起源以及线粒体与叶绿体的起源,更为先进的一点是,这一兼融假说还解释了多细胞生物的起源以及“寒武纪生命大爆炸”现象产生的原因。但这毕竟是一个假说,还需要进一步的验证。
关键词: 原始病毒 内含子 细胞核 内共生 起源 兼融假说 四域分类 WLG团队
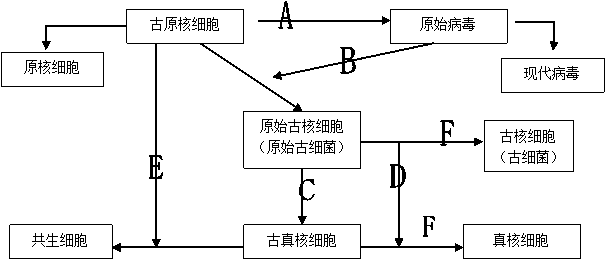
图1 兼融假说图示
注:① A,原始病毒从古原核细胞中逃逸 B,原始病毒侵染古原核细胞
C,质膜包裹DNA形成细胞核 D,原始古核细胞被古真核细胞吞噬
E,古原核细胞被古真核细胞吞噬 F,5次地球生命大灭绝
② 该图仅显示主要生物的进化历程,并不包括所有。
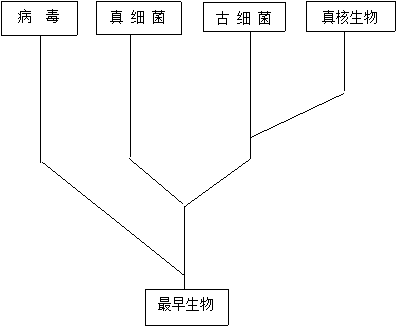
图2 四域分类图示
正文
1 古原核细胞的产生
1952年,美国芝加哥大学研究生米勒在导师尤里指导下,进行了模拟原始大气中雷鸣闪电的实验,获得了20种有机化合物。其后,许多学者模拟原始地球的环境条件,使用不同成分的混合气体、采用不同的能源及选用不同的催化物,成功地进行了多种非生物有机合成模拟实验,获得了包括生物体中常见的20种氨基酸、单羧酸、核糖、脱氧核糖、脂肪酸、核苷酸,以及核苷酸中的碱基,甚至ATP等。由此可知,原始细胞形成的基本条件已经具备,那核酸呢?经过大量的研究发现,少数RNA病毒靠RNA自我复制传递遗传信息;某些RNA也可以在一定条件下充当酶的角色,催化蛋白质合成,还能催化新的RNA合成;核酶对RNA的剪切和短链RNA的聚合也有催化作用。因为RNA有多种功能,所以RNA最有可能是地球上出现的第一批基因和酶。实验也证实核苷酸单体在粘土表面可形成短链RNA分子,新合成的RNA分子还可以与原始RNA分子互补配对。由此可以推测, 最早形成的原始细胞是以RNA为遗传物质的。
但因为RNA的不稳定性,使得以RNA作为遗传物质的原始细胞没有办法在恶劣的原始生存环境中长久的生存下来,取而代之的,先是形成了以双链RNA为遗传物质的原始细胞,接着以RNA为模板,在反转录酶的作用下,合成了单链DNA,形成以了单链DNA为遗传物质的原始细胞,很快,单链DNA又合成了其互补链,形成了双链DNA,这就形成了我们接下来要描述的古原核细胞,即具有双链DNA的古原核细胞(当然,古原核细胞包括以单链RNA、双链RNA和单链DNA为遗传物质的原始细胞,不过,我们下面讨论的主要是以双链DNA为遗传物质的古原核细胞)。从分子生物学角度来看,RNA分子中核糖的C2位上有羟基,较之DNA分子上的脱氧核糖,前者的化学性质很活泼,从而使得RNA链不稳定,再者,双螺旋结构兼具高的键合强度和好的柔韧性,且A=T双键与C三G三键的产生,大大增加了细胞遗传物质的稳定性,即以双链DNA为遗传物质的古原核细胞具有更强的生存优势,这才使得原始细胞得以较好的生存下来。
古原核细胞产生于太古代【太古代,约从38亿年至26亿年前,长达12亿年】,最初可以直接从外界环境得到 ATP,使用存储于 ATP 分子中的化学能来维持自己的生命活动。业已证明,ATP 属于在原始海洋中有可能形成的有机小分子之列。 随着ATP被逐渐增多的原始细胞抢着使用完了,那些能够分解摄入其他有机分子,从中取得能量推动 ATP 合成的原始细胞,即自己在细胞内能合成 ATP 的原始细胞显然具有发展起来的优势。这就是原始的异养式的营养方式 。 再进一步,有些原始细胞中有铁卟啉类分子,取得光合作用成功,能够利用太阳光能来固定 CO2 ,葡萄糖等营养物分子,于是出现了自养式的营养方式,这类古原核生物的典型代表为古原核蓝藻【古太古代,约36亿年前】,人们在非洲南部和澳大利亚西部发现的35亿年前叠层石中就有类似于现今的蓝细菌(或称蓝藻)的球状和丝状的原核生物,光合作用的出现使得大气中氧气的含量不断上升,为后面生物的进化与发展奠定了基础 。
2 原始病毒的产生(伴随性假说)
当时恶劣的生存环境导致古原核细胞(包括之前的以单链RNA、双链RNA和单链DNA为遗传物质的原始细胞)内部基因的表达环境十分不稳定,因而在某一时刻,在细胞正常的生命活动过程中(包括复制、转录、翻译、分裂等),一部分非生命形态的物质(基因与蛋白质的复合体)趁机从古原核细胞中逃了出来,成为了人们所知的原始病毒【太古代,约从38亿年至30亿年前】,即病毒是伴随着细胞的产生而产生,这就是我们在原始病毒的内源性学说的基础上,进一步完善提出的病毒起源的伴随性假说。我们认为,起初绝大多数原始病毒并没有侵染古原核细胞的能力,又因原始病毒不能独立存活,再者又受到周围环境的强烈刺激,病毒基因发生剧烈变异,从而导致原始病毒大量死亡,根据达尔文进化论可知,一小部分本身或变异成了具有侵染性的病毒,侵染当时地球上仅存在古原核细胞,并成功增殖,因而这部分具有侵染能力的病毒才得以存活下来。
支持这一假说的为目前比较流行的病毒起源的内源性学说(即病毒起源于宿主细胞中的RNA或DNA成分的学说),该学说认为病毒来源于正常细胞的核酸,因偶然途径从细胞内脱离出来而变为病毒。因为病毒是专性细胞内寄生的,它们只有在细胞内才能表现出生命现象,脱离细胞后就不能繁殖,因此这样的病毒是不可能在细胞之前起源的;病毒的基因组与其寄主的基因组在结构特点上十分相似,有些甚至在核苷酸序列上与寄主基因组的某些区域也无太大差异;此外,病毒的结构与现代细胞内的核酸和蛋白质的复合体——核蛋白的结构也有相似之处。所有这些事实使得人们现在比较普遍地认为病毒是由细胞衍生的,是细胞内“逃跑”出来的一些基因及蛋白质的复合体。
支持这个学说的多半来自于一些实验的间接证据:病毒与质粒的相似性,质粒本属于细胞的一部分,但它可以随时脱离细胞,并在细胞之间传递;有很多DNA病毒,如细菌病毒中的λ噬菌体,植物病毒中的花椰菜花叶病毒,动物病毒中的乙肝病毒和腺病毒等,这些病毒的DNA或全部或部分可以结合到它们所寄生的细胞的染色体上,从而变为细胞的一部分,这正好是细胞核酸外逸的逆过程;利用核酸分子探针技术发现很多能与细胞染色体结合的病毒DNA的整合区序列与发生整合的染色体的侧翼序列有很大的同源性,尤其在一些逆转录病毒(艾滋病的病原——人类免疫缺陷病毒即属于此类)中的癌基因V-onc与细胞中的原癌基因C-onc高度同源;正常细胞中存在较广泛的逆转录型可动遗传因子,如酵母细胞的Ty因子、果蝇的Copia样因子、脊椎动物的IAf,基因,和逆转录型重复序列,如人的Alu因子及KpnⅠ因子等。另外,在1999年第十一届国际病毒学大会对病毒在基因水平上的起源及进化提供了新的证据:在一种古核生物(一种介于原核生物和真核生物之间的第三类生物)中发现了被称之为“反转子”的遗传单元,它是仅含一个基因并且能自我复制的一段核酸分子,具有重要的基因捕获功能。其基因序列与宿主细胞等位基因高度同源,并且在密码子的使用频率上也有高度相似性,反转子从细菌基因库中捕获基因,扩大自己的遗传信息量,增加生物学功能,最原始的感染性病毒颗粒由此产生,原始病毒采取不同的复制策略,于是产生不同病毒的进化。可见,生命的起源是从细菌开始的,细菌是病毒进化的摇篮。
当然,有的学者认为,病毒是由原始的细胞经过几十亿年的寄生生活,逐渐退化而成的。我们不赞同这一类观点,下面我们拿营寄生生活的支原体、立克次氏体、衣原体(这三类同属革兰氏阴性,代谢能力差,主要营细胞内寄生的小型原核生物,从支原体、立克次氏体至衣原体,其寄生性逐步增强)与病毒相比较(表格1):
| 比较项目 | 支原体 | 立克次氏体 | 衣原体 | 病毒 |
| 细胞构造 | 有 | 有 | 有 | 无 |
| 含核酸类型 | DNA和RNA | DNA和RNA | DNA和RNA | DNA或RNA |
| 核糖体 | 有 | 有 | 有 | 无 |
| 细胞壁 | 无 | 有(含肽聚糖) | 有(不含肽聚糖) | 无 |
| 细胞膜 | 有(含甾醇) | 有(无甾醇) | 有(无甾醇) | 无 |
| 繁殖时个体完整性 | 保持 | 保持 | 保持 | 不保持 |
| 大分子合成能力 | 有 | 有 | 有 | 无 |
| 产ATP系统 | 有 | 有 | 无 | 无 |
| 氧化谷氨酰胺能力 | 有 | 有 | 无 | 无 |
| 对抑制细菌抗生素的反应 | 敏感(对抑制细胞壁合成者例外) | 敏感 | 敏感(青霉素例外) | 有抗性 |
表格1
从表格中我们可以得知,经过长时间的寄生生活,支原体、立克次氏体与衣原体在细胞结构与功能上都有不同程度上的退化,但它们都有共同的特征,即拥有细胞构造,细胞内同时含有DNA与RNA,有核糖体,繁殖时能保持个体完整性,有大分子合成能力,对抑制细菌的抗生素都有一定的敏感性等,相比来说,病毒却没有这些特征,假如说病毒是原始细胞因营寄生生活逐渐退化而来的,很显然,总应该保留一些上述三种寄生性原核生物的共同特征,事实是,一点也没有。另一方面,我们来比较一下线粒体与叶绿体(建立在线粒体和叶绿体内共生起源学说的基础之上),二者也是在细胞内经过几亿年的共生生活,结构和功能上也发生了一定程度的退化,但是线粒体和叶绿体都含有遗传物质DNA和RNA ;线粒体和叶绿体都有膜结构 ;线粒体和叶绿体都能产生ATP ;线粒体和叶绿体都含有核糖体,也能够合成生物大分子;对抑制细菌的抗生素也都有一定的敏感性;二者在繁殖时都能够保持个体的完整性等。以上的这一系列的证据显示,病毒是由细胞退化而来的这一说法是站不住脚的,最起码这些疑问是没办法解答的。
总的来说,病毒最有可能是从古原核细胞中逃出来的一部分非生命形态的物质,经过大自然的筛选,病毒具有了侵染性,才得以生存与增殖开来。
3 内含子的形成(病毒侵染性假说)
随着病毒不断的侵染古原核细胞,一部分病毒基因重组到古原核细胞的基因中,同时也导致了古原核细胞的基因发生突变,致使一部分可表达基因变为沉睡基因(病毒基因与沉睡基因统称为无用基因),再加上其他一些环境因素,导致古原核细胞大量的死亡,但因古原核细胞基因的复制与重组(同时那些无用基因被正常基因分散开来),一部分古原核细胞进化出来了一种应变机制,其中的一些无用基因被古原核细胞重新利用,用以对基因表达的调控,另外一些无用基因则与正常基因同时转录,但在随后的修饰过程中被切除掉,有些无用基因甚至不被转录。久而久之,那些能被转录但不能被翻译的基因成为了如今的内含子。由于内含子的产生,小片段的基因之间可以发生一定程度上的重组,使得基因变异率得到了一定程度上的提升,使得这些拥有内含子的细胞能更好得适应恶劣的环境,这些细胞就是如今我们众所周知的古核细胞(古细菌)的祖先——原始古核细胞(原始古核生物)【时间约为太古代后期,约从30至26亿年前】,这也是我们综合各位学者的观点提出来的病毒侵染性假说。另外,部分原始古核生物演变为了新的物种,但因为没有稳定的内部环境,这些生物并没有得到长足的进化与发展,数量与种类都较少。
古核生物或称古细菌是20世纪80年代出现的名称。古核生物是一些生长在极端特殊环境中的细菌,最早发现的古核生物是产甲烷细菌类,根据其16S rRNA 核苷酸序列的同源性测定分析,它与其他原核细胞相差甚远;相反,其16S rRNA 序列分析和其他一些分子生物学特征却与真核细胞更为近似。后来又陆续发现其他的古核生物,如盐细菌(生长在浓度大的盐水中),热原质体(生长在煤堆中),硫氧化菌(生长在硫磺温泉中)等等,现已发现几百种古核生物。
很多资料显示,古核生物与原核生物相比较,前者与真核生物有较近的亲缘关系,即真核生物很可能是由古核生物的祖先——原始古核生物而不是由古原核生物直接进化而来的。
(1),细胞壁的成分:原核生物的细胞壁主要由含壁酸的肽聚糖构成,古核生物也有细胞壁,但其成分却与真核细胞一样完全不含上述成分,因此抑制壁酸合成的链霉素、抑制肽聚糖前体合成的环丝氨酸、抑制肽聚糖合成的青霉素与万古霉素等对原核生物类有强的抑制生长作用,而对古核生物与真核细胞却无作用。
(2),DNA与基因结构:真核细胞的DNA中含有大量的重复序列,原核生物类的细胞DNA不含重复序列,而古核细胞DNA中却有重复序列存在。此外,多数古核细胞的基因组中像真核细胞一样存在内含子,而原核生物中是不存在内含子的。
(3),核小体结构:古核细胞具有组蛋白,而且能与DNA构建成类似核小体的结构,但与真核细胞典型核小体有差异。
(4),核糖体:众所周知,真核细胞的核糖体为80S,含有70~84种蛋白质,大部分原核生物的核糖体为70S,含有55种蛋白质,而多数古核生物类的核糖体较原核生物有增大趋势,含有60种以上蛋白质,介于真核细胞与原核生物之间。抗生素能作用于70S核糖体,从而对所有原核细胞都能起到抑制蛋白质合成的作用。然而这些抗生素都不能与真核细胞80S核糖体结合而起到对真核细胞蛋白质合成的抑制作用。令人惊讶的是,以上这些抗生素同样不能抑制古核细胞类的核糖体的蛋白质合成。综上所述,古核细胞的核糖体与原核生物的差异很大,从对抗生素的反应看,应更类似真核细胞的核糖体。
(5), 5S rRNA:近20多年来,根据对5S rRNA序列的分析,在鉴定物种进化亲缘关系方面做了很多可贵的工作。古核细胞、原核细胞与真核细胞三者都具有5S rRNA,它们仅含约120个核苷酸。Husysman 与Wachter(1986)根据对5S rRNA 的分子进化分析,认为古核生物与真核生物同属一类,而原核生物却与之相差甚远。5S rRNA二级结构的研究也说明很多古核生物与真核生物相似。
(6),除上述以外,根据对DNA聚合酶分析,氨酰tRNA合成酶的作用、起始氨酰tRNA与肽链延长因子等分析,也提供了类似依据,说明古核生物与真核生物在进化上的关系较原核生物类更为密切。
4 细胞核的形成(或古真核细胞的形成)
真核生物最有可能起源于原始古核生物。因此,我们做出了以下判断:一部分原始古核生物适宜生活在当时恶劣的环境中,因而进化缓慢;另一部分原始古核生物继续进化,先是为了扩大细胞的新陈代谢效率,细胞膜内陷,逐渐的形成了原始内质网【元古宙时期,约从26至22亿年前】。随着进化过程的进行,原始古核细胞需要更为强大的代谢速率以及更为稳定的内部环境,因而一小部分原始古核细胞的原始内质网将遗传物质——基因包裹了起来,形成了原始的细胞核【元古宙后期与古元古代时期,约从22至15亿年前】,最早的真核生物化石发现于大约16~20亿年前的地层中,如加拿大南部的冈弗林特燧石层和我国长城群串岭沟页岩中的化石。起先,原始细胞核膜上有很多孔洞(如单细胞真核生物,双滴虫类),并没有起到隔绝的效果,但相对于原来已是有了很大的进步,一段时间后,这类原始细胞核就进化出来了核孔,有了核孔,细胞核内外的物质交换就得到了一个有效的调控,因而细胞核内便形成了一个相对稳定的基因表达环境。此时的原始古核细胞被称古真核细胞【古元古代与中元古代,约从15至8.5亿年前】。
4.1 原始核膜的形成(非内共生起源学说)
这种观点能比较好地说明现代真核细胞中核膜和内质网的连续性。核膜在细胞的有丝分裂过程中消失,分裂后参与核膜重建的一般除了有原来核膜的碎裂物外,还有内质网的成分。因此,核膜和内质网实际上是同一类膜系统,甚至可以认为核膜是内质网的一个特殊的组成部分。那么,双层的核膜又是怎样产生的呢?
这可以从内质网的结构特点来说明:内质网也往往是双层膜结构,两层膜之间的距离与核膜内外膜的间隙差不多。核膜在细胞的进化过程中正是起源于具有双层膜结构的原始内质网。把原始内质网作为核膜的前身,而原始内质网又是起源于细胞膜,所以核膜不是直接源于细胞膜,而是通过原始内质网过渡而来。这种观点能较好地说明核孔的形成,因为可以认为首先是具有双层膜结构的原始内质网的两层膜在某些区域融合而产生穿过两层膜的孔道,然后再由有了这种结构的原始内质网形成核膜。从而,这种原始的核膜并不是完全封闭的,而是有很多能与细胞质畅通无阻的孔道,这些孔道就是原始的核孔。原始核孔以后再进化发展为能控制细胞核与细胞质物质交换的核孔。这样的一种过程并不存在刚形成核膜时细胞核与细胞质之间难以进行物质交换的问题。实际上,现代具双层膜结构的内质网系统确是有不少贯通双层膜且形状不规则的孔道,其中一些孔道的大小还和核孔差不多。因此,原始的内质网同样是可有这类孔道的,这就使得原始的核膜也具有这类孔道。这些孔道是原始的细胞核与细胞质沟通的桥梁,也是形成真正的核孔所必需的结构基础。
依照这种看法,原始的核膜上是有很多完全开放的孔道,那么现在是否能找到这种原始核膜的遗迹呢?最近发现,一些十分低等的单细胞真核生物(如双滴虫类)的细胞核的核膜有不少大小不一的缺口,而在这些核膜上却没有观察到核孔的存在,从而这些生物的核膜很可能就是原始核膜的遗迹。这一事实对上面这种关于核膜与核孔起源的看法是一个有力的支持。至于现代核膜的内外膜在形态结构与化学组成上的差异,则可以认为是在核膜起源后再分化产生的。
综合起来,这一种观点是核膜起源,也就是细胞核起源的最有可能的方式和途径。另一方面,核膜是双层膜还有其内在的原因,这一原因就是核膜必须有大口径的核孔才能满足细胞核与细胞质之间的物质交换。如果在单层膜上,即使可以产生如此大的孔,也是一种不稳定的结构,只有在双层膜上,通过两层膜的部分融合,才有可能形成大口径且稳定的孔道。
细胞核的产生使得真核细胞的核质相对分离,遗传信息的转录与翻译分别在核内和细胞质中进行,而在古原核细胞中这两种过程是可以同时同地进行的。细胞核的建立还提供了一种有利于其内的基因组向复杂化、多功能化发展的环境。由于基因组的复杂化与多功能化,使得遗传信息信息量大大增加,继而一个核所能控制的细胞质的量也大大增加(一个细胞核内所含的遗传信息量与其所控制的细胞质的量呈正相关,但一个核内所含的遗传信息量是有一定限度的,因而其所控制的细胞质的量也是有一定限度的),这就是为什么如今真核细胞(大多数较大,10~100微米)一般比原核细胞(大多数很小,1~10微米)大。
另一方面,由于细胞核的产生,使得基因有了一个比较稳定表达环境,再者遗传信息量的增加与基因的变异,为古真核细胞进行有性生殖奠定了一个坚实的基础,又因为有性生殖相比无性生殖有更大的生存优势,经过环境的不断筛选,有部分古真核生物进化出来了有性生殖。在如今的单细胞原生动物中,有很多种类既可以以分裂或出芽方式进行无性生殖,也可以以减数分裂产生配子进行有性生殖。如可引发人类和其他动物感染的疟原虫,可引起人类睡眠的锥虫等,有性生殖的出现使得生物的进化得到了一次飞跃,生物种类的数目得到了一定程度上的增加。
4.2 多细胞生物与有性生殖的出现
紧接着,也就进入了震旦纪时期【震旦纪时期,约从8.5至5.7亿年前】,由于细胞核的出现,遗传信息量不断增加,遗传信息表达的稳定性不断得到增强,使得细胞得以进行充足的发展,逐渐形成了多细胞生物。从化石记录看,多细胞真核生物大约出现在6亿年前,在我国贵州中部的磷块岩中发现两种类型植物的化石(大约6亿~6.5亿年前):一种可能是细胞集群;第二种是细胞分化明显的叶状体植物(叶藻),其内部有皮层和髓部的分化,髓部还有薄壁组织和假薄壁组织的分化。多细胞生物体的出现使得生物体内部的细胞出现了分工,出现了细胞器官的分化,不同功能的细胞的形态也开始分化,另一方面,由于细胞器官的分化,使得细胞具有了比较完善的生殖系统,有性生殖的完善,也使得生物的变异速率大大提升,出现了多种多细胞生物。现存的团藻可能反映出较早出现的多细胞生物的某些特征。团藻实际上是单个细胞经过多次分裂,后代仍聚在一团未曾分开而成的个体,但是在这团细胞中,已有一定的分工,有的细胞特化其运动功能,有的细胞特化其光合作用制造食物功能,有的细胞特化其有性生殖功能。粘细菌通常以单细胞阿米巴方式活动,以吞噬细菌为生,在遇到饥饿的情况下,单个细胞聚集起来,形成蛞蝓状的多细胞个体,并能进一步分化为基部、孢子梗、孢子囊等几个部分。孢子囊中每个孢子在适宜条件下可发展成一个阿米巴单细胞个体。极少数多细胞生物可以进行有氧呼吸,因而可以形成一些体积稍大的生物,但因细胞自身代谢速率的限制,绝大多数只能形成一些体积比较小的多细胞生物。
5 叶绿体与线粒体的由来(内共生学说)
古真核细胞继续进化,由于细胞核的出现与其功能的不断完善,再加上细胞内部各种结构的优化,古真核细胞有了更强的吞噬与调控能力,一部分古真核细胞吞噬了原始古核细胞(原始古核蓝藻或原始古核细菌)【时间约在震旦期末期,寒武纪早期,5.4亿年前】,并很好的调控它们,为己所用。由于两者亲缘关系比较相近,再者两者能够互利共生,因而原始古核细胞在古真核细胞中生长繁殖开来,慢慢地,原始古核蓝藻演变为了叶绿体,原始古核细菌演变为了线粒体(蓝小体是一种进化中的古核蓝藻,即介于古核蓝藻与叶绿体中间)。当然,一部分古核细胞吞噬了古原核细胞(或原核细胞),继而发展成为了如今的共生细胞(如池沼多核变形虫细胞中的共生细菌),但因两者亲缘关系相差较大,不能十分有效的合作,故而这些古原核细胞(或原核细胞)并没有演变成为线粒体或叶绿体。
5.1 内共生学说
支持这一论断的学说为内共生起源学说,所谓内共生就是根据自然界中比较普遍地存在着细胞内共生这一现象,认为一些细胞器也是通过细胞内共生的过程而起源的。设想一种较大的单细胞生物“吞吃”了另一种较小的单细胞生物,两者首先建立起内共生的关系,然后在细胞进化的过程中,被“吞吃”的小细胞逐步高度特化,不能再在细胞外长期生存,从而就成为了细胞内的一种具有专门功能的细胞器。
内共生的例子在现代生物界还是有不少的。有真核细胞共生于其他种类的真核细胞的情况,如许多种单细胞的绿藻、甲藻和硅藻可以共生于高等植物、真菌、其他藻类以及脊椎动物和无脊椎动物的细胞中。此外,也有原核细胞共生于真核细胞的情况。例如,蓝藻可以共生在真菌、变形虫、鞭毛虫以及已失去叶绿体的绿藻的细胞中,蓝藻可以进行光合作用,为宿主提供一定的养分;细菌也可以共生在真核生物的细胞中,在不少昆虫的特殊细胞中,就有正常共生的细菌,这些细菌对于宿主来说往往是有重要生理意义的;特别有意思的是,一种巨大的(大小有2至3毫米)池沼多核变形虫的细胞中并没有线粒体,但却有一些需氧的胞内共生细菌,这些细菌实际上是起到了线粒体的作用。
另外,一些学者在这方面也做了一些实验。在60年代中期,有人将细菌感染大变形虫,新感染细菌的变形虫其细胞生命活动受到一定程度的有害影响。但经数年维持培养后,其中细菌对变形虫的有害作用逐渐变小,最终,变形虫须依赖所获得的细菌才能生存下来。后来又有人以人工诱导变形虫吞噬作用或以显微注射方法将外来细菌引入到含有细菌共生体的变形虫细胞内,观察到,细菌由变形虫质膜包裹形成吞噬体,经历一个消化过程,其中少数吞噬体能避免被消化而成为包有共生细菌的小泡。实验后4~10天,约有5%的细菌以这种形式存活下来,在变形虫细胞内继续繁殖。其实,支持内共生学说的依据还有很多:
(1),现代真核细胞中的线粒体或叶绿体只能由原有的线粒体或叶绿体通过分裂、断裂或出芽等产生,而不能从细胞的任何其他部分重新形成。失去线粒体或叶绿体的真核细胞是不能再产生这些细胞器官的。线粒体与叶绿体这种分裂繁殖的方式与细菌的繁殖方式类似。
(2),把线粒体从一种真核细胞中分离出来后再引入到另外一种真核细胞中,便可以观察到这些异源的线粒体能在细胞中生存很长的时间。叶绿体在异种细胞中的生存能力就更大了。一些海螺靠吸食海藻为生,在它们的消化细胞中可以发现所食那种海藻的叶绿体。这些叶绿体不但能很好地生存,而且在光照下仍然可以进行光合作用,从而有叶绿体的海螺在光照下即使长期不进食也不会饿死。这些海螺的卵和幼虫内都没有叶绿体,表明叶绿体是在海螺进食海藻后才从海藻细胞进入其细胞内的。此外,离开细胞的线粒体和叶绿体都有一定的独立生存能力,叶绿体甚至可以在细胞外人工培养。人工培养的叶绿体可进行一定次数的分裂,并在相当长的时间内保持光合作用的能力。
(3),在某些现代的真核细胞中,可以找到一些介于胞内共生蓝藻与叶绿体之间的生物结构。例如在一种真核藻类的细胞内,只有称为蓝小体的东西而没有通常所说的叶绿体。蓝小体跟蓝藻特别相似,以至曾有人把它们作为一种生物那样命名。但后来发现,蓝小体并不是一种蓝藻,它们的DNA已和叶绿体的基因组差不多。
(4),从显微形态结构、化学成分、蛋白质来源等方面的差异可以看出,线粒体和叶绿体的两层膜是有不同的进化来源的。如外膜与细胞的内膜系统相似,可与内质网与高尔基体膜沟通;内膜与细菌质膜相似,内膜内折,分别形成细菌的间体、线粒体的嵴和叶绿体的类囊体,他们的电子传递均表现出与膜结合在一起的特点;存在于线粒体或叶绿体和基质中特有的有氧呼吸(或光合作用)的酶系,也发现存在于细菌质膜和基质中。在膜的化学成分上,内膜的蛋白质与脂质的比值远大于外膜,因此内膜更接近于细菌质膜的成分。给离体线粒体标记放射性氨基酸,只有内膜被标记,说明其内膜的蛋白质成分一部分是自己合成的。
(5),线粒体和叶绿体都有自己特有的基因组,这些基因组在大小、形态和结构方面与细胞核的基因组有较大的差异,而却与细菌的基因组比较相似。特别的是,这两种细胞器官中DNA复制与RNA转录所需的酶都与一般细菌中相应的酶相似,且能独立复制和转录自己特有的RNA,其mRNA、rRNA的沉降系数均与细菌的类似。
(6),线粒体和叶绿体中蛋白质合成的体系,包括核糖体(rRNA和核糖体中的蛋白质)、mRNA、tRNA以及有关的酶,还有对蛋白质合成抑制物的反应,都与一般细菌中的相似,而却与真核生物细胞质中的有较大差异。
(7),近年来分子进化的有关研究表明,线粒体中的rRNA序列与一般细菌中的比较相似,而与线粒体所在的细胞质中的rRNA序列则有较大差别。这是从分子水平上对线粒体内共生起源学说的一个有力的支持。对其他生物大分子(包括一些蛋白质)的序列分析也得出相似的结论。通过这些分析,还肯定了线粒体的祖先与现代称为紫细菌的那一个类群中的某些种类相似。在叶绿体方面,叶绿体rRNA的序列与蓝藻中的rRNA序列的相似程度很高,而与叶绿体所在的细胞质中的相应序列却相差较大。分子进化的研究还进一步确定了叶绿体是起源于现代某一类蓝藻的远古祖先。
(8),由内共生学说可以推出这么一条结论,即生物界里很可能会存在这样四类真核微生物:既没叶绿体又没线粒体,没叶绿体但有线粒体,有叶绿体但没线粒体,有叶绿体又有线粒体。而事实是,在地球上,这四类真核生物真的存在,它们的典型代表依次是贾第虫、高等动物细胞、眼虫、高等植物的叶肉细胞。这进一步证明了在叶绿体与线粒体起源方面,内共生学说的正确性。
5.2 寒武纪生命大爆炸
生命离不开适当的温度,生物最适合的温度范围在0~45Cº之间,在寒武纪之前,地球上出现过长达几千万年的冰期,冰雪把整个地球包裹起来,厚厚的冰雪把绝大部分日光反射回宇宙空间,地球表面温度一直在零下50Cº左右,当时的生物只能在低温中勉强度日。在寒武纪早期,冰雪融化,地表开始升温,赤道附近的海洋水温达20Cº以上,非常适合生物大规模生长;另外,经过海洋藻类几十亿年的光合作用,生产出越来越多的氧气,到寒武纪初期,大气中氧气含量已达10%,约为现代氧气含量的一半;当然还有海洋中磷元素的增加,这些都是外因。
内因更为重要,由于线粒体与叶绿体的出现(当时还不能称之为线粒体与叶绿体)与 大气中氧的增加,使得大多数生物体有能力并开始利用氧气使有机物氧化分解,由此所得的能量比以往无氧条件下分解所得的能量多几十倍,大量获得能量,显然极大的提高生命活动效率,使得细胞有能力支撑一个大型的有机体,因而大量的大型多细胞生物出现,这就进一步促进了器官的分化,极大地完善了多细胞生物的有性生殖系统,从而更进一步地促进了多细胞生物的进化。这一时期,也正好发生在寒武纪的早期,因而在在寒武纪开始后的短短数百万年时间里,包括现生动物几乎所有类群祖先在内的大量多细胞生物突然出现,这一爆发式的生物演化事件被称为“寒武纪生命大爆炸”。寒武纪是现代生物的开始阶段,是地球上现代生命开始出现、发展的时期。 寒武纪常被称为“三叶虫的时代”,这是因为寒武纪岩石中保存有比其他类群丰富的矿化的三叶虫硬壳。寒武纪是显生物的开始,标志着地球生物演化史新的一幕。带壳、具骨骼的海洋无脊椎动物趋向繁荣,它们营底栖生活,以微小的海藻和有机质颗粒为食物,其中,最繁盛的是节肢动物三叶虫,故寒武纪又称为“三叶虫时代”,其次是腕足动物、古杯动物、棘皮动物和腹足动物,寒武纪的生物形态奇特,和我们现在地球上所能看见的生物极不相同。
很快,当细胞内的遗传信息量、细胞的新陈代谢速率与多细胞生物体的体积达到了一个平衡,生物的进化速率就降了下来,各种生物的性状也就开始稳定下来,各种生物也就开始了它们缓慢而又漫长的进化。另一方面,大气中已积累起臭氧层,把来自太阳辐射中的紫外线吸收掉 90% 以上。由于紫外线对生物大分子,尤其是核酸大分子有很大破坏力,减少紫外线辐射,可极大的保护生物免遭遗传物质破坏而导致的突变和死亡。由此,生物细胞有了更长的寿命和更强的稳定性。这就是为什么现今世界上几乎所有的动物的门都在这一时期同时出现,而之后再没有产生新的门的原因。
5.3 5次地球生命大灭绝
古生物化石记录表明,在过去的5.4亿年的地质历史中,地球上总共发生5次生物大灭绝事件,其中距今2.5亿年之前二叠纪末的生命灭绝事件是生命发展史上最惨烈的事件,造成了多达95%的海洋生物和70%的陆生脊椎动物的灭绝。,总之一句话,就是地球上的环境发生了剧烈的变化,生物的生存环境变得尤为恶劣,适合生物生存的空间变得尤为狭窄,大量生物因而灭绝,但有一些生物却能顽强的存活下来。其中不乏有那些能产生抗生素和毒素等物质的生物,它们为了生存,必须要扩大自身的生存空间,不适宜它们生长的环境促使它们产生大量的抗生素与毒素,以此来抑制周围生物的生长,保证自身最基本的生存空间。
经过5次大的抗生素与毒素洗礼之后,很多对抗生素以及毒素敏感的生物大量灭绝,一些原本对抗生素以及毒素敏感的生物在激烈的竞争中,进化出来了对抗生素和毒素不敏感的生物类型。众所周知,抗生素(antibiotics)是由微生物(包括细菌、真菌、放线菌属)或高等动植物在生活过程中所产生的具有抗病原体或其它活性的一类次级代谢产物,能干扰其他生物正常功能的化学物质,这些微生物又是广泛分布在世界的各个角落,这必然为全世界大范围的洗礼奠定了坚实的基础,当然,这一洗礼现象并不是每一个角落都会发生。
在我们的兼融假说里,古原核生物、原始古核生物、古真核生物以及原核生物中绝大多数都是对抗生素以及毒素敏感,经过几次大的洗礼之后,一些种类的生物大量灭绝,一些种类的生物的生存范围急剧缩小,一些种类的生物顽强的生存了下来。
总结:
我们的这一假说(即兼融假说,包括病毒起源的伴随性假说、病毒侵染性假说等)可以很好的说明生物是如何由古原核细胞进化到如今的真核细胞,并能很好的解释在进化过程中遗留的一些问题。
1,其中详细的说明了真核细胞中内含子的由来(包括真核细胞中细胞核、叶绿体、线粒体中的内含子的由来),即是由病毒侵染而来的,继而形成了原始古核生物,在经过几十亿年的进化,成为了如今人们仍可见到的古核生物,这也是为什么古核生物的基因中会含有大量的内含子,与真核生物在很多方面又极为相像。
2,这一假说吸取了内共生起源学说与非内共生起源学说双方的优点,即线粒体与叶绿体是由古核细菌与古核蓝藻在古真核细胞中共存并逐渐演化而来的,而细胞核则是由内质网包裹古核细胞的基因而产生的,从而很好的弥补了上述两种假说的不足之处,即形象的解释了为什么线粒体与叶绿体的DNA含有内含子,但其很多性状却与当今的细菌与蓝藻相似等问题。
3,有细胞就会有病毒,病毒是伴随着细胞的产生而产生,而且新型病毒的产生主要有两个来源,一是在原病毒基因的基础上经变异转化而来,另一部分是从细胞中重新逃逸出来,并经过环境的选择逐渐进化而成。这也能解释某些真核病毒的基因组中也含有内含子,并且这些病毒的基因转录生成成熟的mRNA亦需要类似于真核生物mRNA的拼接过程(去除内含子,连结外显子),而退化性学说却没有充足的证据证明这一现象产生的原因。
4,这一假说又能很好的解释古原核细胞、原始病毒、原始古核细胞、古真核细胞与真核细胞产生的先后顺序,即先产生了古原核细胞,原始病毒随即伴随着古原核细胞的产生而产生,原始病毒侵染古原核细胞,经过很长一段时间的作用,古原核细胞内部产生了内含子,形成了原始古核细胞,原始古核细胞因为自身发展的需要,逐渐形成了原始内质网(细胞膜内陷),原始内质网膜又将含有内含子的遗传基因包裹,形成了原始细胞核,不久形成了核孔,成为了如今的细胞核,因为细胞有了细胞核,赋予了细胞核内的基因一个稳定的表达环境,促使细胞核内的基因不断复杂化、丰富化,使得古真核细胞有能力吞噬较大的物质,并能很好的调控这些物质(这也就是为什么如今发现的大多数含有共生细菌或蓝藻的细胞均为真核细胞的原因),那时原始古核蓝藻与原始古核细菌就被古真核细胞吞噬了进来,因为双方之间可以很好的互利共生,所以原始古核蓝藻与原始古核细菌就在其内部繁殖并生长起来,慢慢的就变成了如今的叶绿体与线粒体,接下来就形成了如今的真核细胞。
5,这一假说又能很好的解释在远古时代生命进化的历程。
在太古代后期,即在30~26亿年前,由于内含子的出现,古核细胞的基因变异率得到了提高,细胞得到了一定程度上的进化,出现了很多新的物种,但因为没有稳定的内部环境,这类生物并没有的到充足的发展。
在15~8.5亿年前,即古元古代与中元古代,此时的古核细胞被称古真核细胞。,由于细胞核的产生,使得细胞得以进行充足的发展,逐渐形成了多细胞生物,但因细胞自身代谢速率的限制,大多只能形成一些体积比较小的多细胞生物。
在震旦期末期,寒武纪早期,约5.4亿年前,古真核细胞继续进化,一部分古真核细胞吞噬了原始古核细胞(原始古核蓝藻或原始古核细菌),由于两者亲缘关系比较相近,再者两者能够互利共生,因而原始古核蓝藻逐渐演变为了叶绿体,原始古核细菌逐渐演变为了线粒体,一方面,由于地球水体温度的升高,温度变得适宜生长与繁殖,另一方面,由于线粒体与叶绿体的出现以及空气中氧气含量的增加,使得细胞的新陈代谢速率得到飞一般的提升,再加上有性生殖系统的不断完善,大量的大型多细胞生物出现,大型多细胞生物的出现反过来使得生物体内各器官的分化更为彻底,这就极大地促进了生物的有氧呼吸能力和代谢效率的提高以及生物体有性生殖的完善,这也就更进一步地促进了生物的进化。因而在在寒武纪开始后的短短数百万年时间里,包括现生动物几乎所有类群祖先在内的大量多细胞生物突然出现,这一爆发式的生物演化事件被称为“寒武纪生命大爆炸”。
6,这一假说也能很好的解释原始古核生物、线粒体与叶绿体都对抗生素敏感,而古核生物却对抗生素不敏感。
即原始古核生物与古真核生物本身是对抗生素敏感的,但经历过5次大的抗生素洗礼过后,原始古核生物与古真核生物大量死亡,但一部分对抗生素不敏感的原始古核生物与古真核生物存活了下来,并得以繁衍。被古真核生物吞噬的原始古核生物因有古真核细胞的保护,因而对抗生素敏感的性质被保留了下来,也就是说,由原始古核蓝藻与原始古核细菌演变成为的叶绿体与线粒体仍是对抗生素敏感的。存留下来的原始古核生物变为了现在的古核生物,吞噬了原始古核细胞的古真核生物就演变为了现在的真核生物。当然,对抗生素敏感的性状在真核细胞生物中并没有完全消失,有一部分真核生物仍然保留着对抗生素敏感的特性,例如人类。
7,兼融假说也是我们生物界四域分类学说的基础,四域分类学说进一步明确了病毒的地位。
我们团队对兼融假说进行了进一步的探究,并大胆的将病毒纳入到生物界里面,独立形成一个域,即病毒域,进而提出了我们的生物界四域分类学说,即在生物进化的早期,存在一类各生物的共同祖先——古原核细胞,由它分出四条路线,最初先分成两支,一支为病毒域(这一条途径比较特殊,因为病毒并不是由古原核细胞进化而来的,而是从古原核细胞中逃逸出来的一部分非生命形态的物质),一支为真细菌——古细菌——真核生物域,后者进一步分为真细菌域和古细菌——真核生物域,古细菌——真核生物域最终分为古细菌域与真核生物域。即将自然界的整个生物重新划归为四大域,真核生物域、古细菌域、真细菌域与病毒域。将非生命形态的病毒与生命形态的细胞联系起来,使得生物分类更加的完善。
如果这一假说成立,那么,生物进化领域内的很多问题就能够迎刃而解,当然,由于实验条件的限制,这一假说中还存在很多问题,这需要日后进一步的研究与探讨。
参 考 文 献
【1】吴相钰,陈守良,葛明德主编 . 陈阅增普通生物学 . 第3版 . 北京:高等教育出版社,2009
【2】谢天恩,胡志红主编 . 普通病毒学 . 第1版 . 北京:科学出版社,2002
【3】周德庆 . 微生物学教程 . 第2版 . 北京:高等教育出版社,2002
【4】瞿中和,王喜忠,丁明孝主编 . 细胞生物学 . 第3版 . 北京:高等教育出版社,2007